

APA Reference
Mashrafi, M. (2026). Universal Life Competency- Ability Framework and Equation: A Conceptual Systems-Biology Model. International Journal of Research, 13(1), 92-109. https://doi.org/10.26643/eduindex/ijr/2026/5
Mokhdum Mashrafi (Mehadi Laja)
Research Associate, Track2Training, India
Researcher from Bangladesh
ORCID ID: https://orcid.org/0009-0002-1801-1130
Abstract
Living organisms across biological taxa—including humans, animals, birds, fish, insects, plants, and microorganisms—can be conceptualized as open thermodynamic systems that sustain internal order through continuous exchange of matter and energy with their environments. While extensive work in physiology, ecology, and systems biology has investigated metabolic scaling, resource assimilation, and energy budgets, few integrative frameworks exist for synthesizing absorption processes, physiological losses, organismal mass, and biochemical competency into a unified comparative model that applies across taxa.
This paper presents the Universal Life Competency–Ability Framework, a conceptual systems-biology model that formalizes biological performance as the product of three core determinants: organism mass (M), net resource uptake rate (AE − TE), and a composite competency coefficient (CE) capturing biochemical and physiological efficiency. The resulting index is not proposed as a physical law but as a scalable, mass-balance-based metric that enables comparative interpretation of biological performance across life forms.
The Introduction reviews existing biological models related to mass-energy balance, metabolic scaling, ecological energetics, and plant–animal physiology, highlighting the conceptual gap addressed by this framework. The Methods section derives the model using established thermodynamic and physiological principles and defines all parameters. Results demonstrate how the model applies across major taxa through conceptual scenarios rather than numerical predictions. The Discussion interprets biological implications, examines alignment with existing theories, and identifies limitations and future research directions.
Findings suggest that organisms experiencing positive net uptake (AE > TE) and high competency (CE) exhibit greater biological performance and resilience, while those in nutrient deficiency, disease, or stress states exhibit reduced net uptake and diminished competency. Importantly, the framework aligns with empirical observations in plant physiology (photosynthesis–respiration balance), animal nutrition (intake–expenditure models), and ecological energetics (net primary productivity and trophic transfer).
This systems-level model offers a unifying conceptual lens for interpreting cross-taxonomic variation in growth, vitality, and function without overclaiming precision or universality. It complements existing detailed models by emphasizing emergent principles shared across living organisms. Future work may formalize empirical estimation of CE, integrate species-specific scaling exponents, and explore applications in agriculture, environmental physiology, conservation biology, and bioengineering.
Keywords: systems biology, mass balance, bioenergetics, metabolic scaling, competency, physiology, open systems, life processes.
1. Introduction
1.1. Background
All living organisms operate as open, nonequilibrium thermodynamic systems, continuously exchanging matter and energy with their surroundings in order to sustain their biological structure and function. In contrast to closed or isolated systems, living organisms cannot maintain internal order without importing resources and exporting waste; their survival depends on a constant flow of substrates through metabolic networks. From the perspective of classical and statistical thermodynamics, life represents a persistent reduction of local entropy, achieved by importing low-entropy inputs—such as nutrients, light, water, and oxygen—and exporting high-entropy waste outputs, including heat, carbon dioxide, and nitrogenous compounds (Schrödinger, 1944; Nelson & Cox, 2021). By doing so, organisms counteract the natural tendency toward disorder and sustain the chemical disequilibria necessary for molecular self-organisation, signalling processes, and biosynthesis.
This continuous energy–matter exchange is not merely a biochemical curiosity; it is the core mechanism underpinning all biological performance. At the cellular level, imported substrates are metabolized to produce ATP, reducing power, and precursor metabolites that fuel anabolic pathways, maintain membrane potential, enable motility, and regulate homeostasis. At the organismal level, these molecular events scale up to support development, growth, tissue repair, immune function, reproduction, and behavioral interactions. Losses in energy or matter—due to starvation, thermal stress, disease, or environmental fluctuations—directly translate into reduced performance, diminished competency, and ultimately mortality if sustained.
Despite the substantial diversity found across the tree of life, living organisms share several systemic properties that are universally required for their persistence. These include: (a) mass-energy throughput, the rate at which organisms assimilate and dissipate resources; (b) metabolic conversion efficiencies, which determine the proportion of absorbed substrates that are converted into usable biochemical forms; (c) physiological losses, such as respiration, transpiration, and excretion; (d) environmentally mediated performance, reflecting how temperature, oxygen availability, light, and nutrient supply modulate metabolic fluxes; and (e) biochemical competency thresholds, referring to the minimal enzymatic, hormonal, and structural integrity required for efficient metabolism. Although expressed differently in plants, animals, microbes, and fungi, these properties define the constraints under which all living systems operate.
Three major scientific disciplines have independently explored these principles. Physiology and biochemistry investigate cellular and molecular mechanisms, including enzymatic catalysis, respiratory pathways, and hormonal regulation, providing insight into the mechanistic basis of metabolism in plants, animals, and microorganisms. Ecological energetics examines how energy and biomass flow through populations, communities, and ecosystems, linking individual physiology to trophic interactions, carrying capacity, and ecosystem productivity. Systems biology, in contrast, focuses on multiscale modeling of networks and emergent behaviors, integrating molecular interactions with organismal phenotypes through computational and theoretical approaches.
Despite substantial progress in each field, there remains no broadly applicable conceptual framework for comparing how diverse organisms assimilate, retain, and convert resources into biological performance outcomes across taxa. Existing frameworks tend to be taxon-specific—photosynthesis models for plants, energetic balance models for animals, and growth kinetics for microbes—making cross-system comparisons difficult. A unifying conceptual perspective is therefore needed to bridge these domains and enable integrated interpretation of biological performance across the breadth of living systems.
1.2. The Problem and Knowledge Gap
Despite extensive scientific progress in physiology, ecology, and bioenergetics, existing quantitative frameworks used to evaluate biological performance tend to be domain-specific rather than integrative. For instance, in plants, performance is commonly assessed using the photosynthesis–respiration balance, which captures the net gain of assimilated carbon after accounting for respiratory losses (Taiz et al., 2015). In animals, biological performance is frequently modeled through dietary intake versus energy expenditure, a framework rooted in nutritional physiology and metabolic energetics (Blaxter, 1989). At the ecosystem scale, productivity is evaluated through net primary production (NPP) and trophic transfer efficiency, which quantify biomass accumulation and energy flow among trophic levels (Odum, 1971). Each of these models is robust within its own domain and has yielded significant empirical insight.
However, while such frameworks excel within specific biological contexts, they lack a unifying abstraction capable of representing biological performance across multiple taxa using shared principles. Specifically, there is no widely accepted framework that simultaneously: (1) spans the diversity of living organisms without relying on species-specific formulations; (2) integrates mass uptake, physiological losses, and biochemical competency into a single cohesive structure; and (3) maintains biological interpretability without overextending into unjustified claims of universal physical laws. This conceptual gap restricts our ability to compare biological performance across plants, animals, microbes, and other life forms in a standardized manner, despite the fact that all rely on similar thermodynamic and metabolic principles.
The persistence of this gap can be attributed, in part, to methodological fragmentation among subdisciplines. Plant biology focuses on carbon assimilation, water relations, and photophysiology; animal physiology emphasizes nutrient intake, respiration, and metabolic demand; microbial metabolism employs substrate kinetics and maintenance energy models; and ecological energetics scales these principles to populations and ecosystems. Although these fields each investigate aspects of mass-energy flux and metabolic efficiency, they rarely converge on a shared analytic language. Consequently, cross-taxonomic comparison of performance metrics—such as productivity, stress tolerance, or vitality—remains conceptually challenging, even though the underlying physiological processes are structurally analogous. Developing a framework that bridges these disciplinary divides would therefore enable deeper comparative insights into the general principles governing life across biological scales..
1.3. Toward a Systems-Biological Perspective
Systems biology provides an integrative framework for understanding living organisms by conceptualizing them as networks of interacting components governed by resource availability, metabolic pathways, and environmental constraints (Kitano, 2002). Rather than treating biological processes in isolation, this perspective emphasizes the emergent properties that arise from coordinated interactions among cellular, physiological, and ecological subsystems. Within this paradigm, biological performance can be understood as the result of dynamic interplay between three key dimensions: resource absorption (inputs), physiological maintenance and losses (outputs), and biochemical conversion efficiencies (internal functional states). Each of these dimensions influences how effectively organisms acquire, retain, and utilize matter and energy to support growth, reproduction, and homeostasis.
This systems-level view aligns closely with mass-balance principles commonly employed in chemical engineering, ecology, and biophysics. In these fields, the governing relationship is often expressed in the generic form:
Accumulation = Inputs − Outputs − Maintenance Losses.
Such formulations capture the fact that net change in biomass or energy content depends not only on acquisition but also on respiratory, excretory, and maintenance costs. Comparable mass-balance expressions appear across a diverse set of biological modeling traditions, including photosynthesis–respiration models in plants, metabolic flux analyses in cellular systems, microbial growth kinetics in chemostat studies, animal energy budget models, and biomass allocation models used in ecology and forestry. Although developed in different disciplinary contexts, these frameworks share the core assumption that biological performance can be quantified through net fluxes of matter and energy modulated by organismal constraints.
The Universal Life Competency–Ability Framework builds upon this mass-balance logic by proposing a biologically interpretable measure of performance grounded in three constituent components: organismal mass (M), which serves as a scaling factor reflecting total metabolic demand; net resource uptake (AE − TE), representing the balance between assimilated and lost substrates; and the competency coefficient (CE), which integrates biochemical and physiological efficiency. Together, these components yield a composite expression for biological performance that captures both the magnitude of resource flow and the quality of internal biological processing.
Importantly, unlike physical energy equations derived from thermodynamic laws, this framework does not claim to produce outputs in joules or watts. Instead, it yields a comparative biological performance index, allowing meaningful interpretation of growth potential, stress resilience, or physiological vitality across taxa without asserting universal physical dimensionality. In this way, the systems-biological perspective provides conceptual grounding for a unifying framework that integrates mass balance, metabolic efficiency, and biochemical competency within a single cross-taxonomic interpretive structure.
1.4. Objective and Scientific Contribution
The purpose of this paper is not to introduce a new universal physical law or to redefine thermodynamic principles, but rather to advance a conceptual systems-biology framework that enables integrated interpretation of biological performance across diverse taxa. Specifically, the framework aims to synthesize mass and energy throughput, unify absorption and loss processes, incorporate physiological and biochemical competency, facilitate comparison among organisms, and connect empirical observations from multiple scientific domains. By doing so, it seeks to address a gap in current biological modeling, where existing approaches are often constrained to particular organisms, metabolic pathways, or ecological contexts.
Within this scope, the central research question guiding this work can be articulated as follows:
How can organismal performance be conceptually modeled as a function of mass, net resource uptake, and biochemical competency in a manner that is scientifically grounded and taxonomically general?
This question reflects the need for a cross-domain framework capable of reconciling diverse empirical findings without relying on species-specific equations or overextending claims into physical universality.
In response to this inquiry, the paper makes four principal scientific contributions. First, it proposes a generalizable, mass-balance-based model that applies to living organisms regardless of taxonomic group, metabolic strategy, or ecological niche. Second, it introduces a competency coefficient (CE) that encapsulates biochemical and physiological efficiency, including enzymatic activity, nutrient sufficiency, hormonal regulation, and tissue integrity. Third, it provides a conceptual bridge between plant and animal physiology, enabling shared interpretation of processes such as photosynthetic assimilation, dietary intake, respiration, transpiration, and excretion. Fourth, it offers a framework for interpreting growth, vitality, and stress in terms of the interplay between mass, net resource uptake, and biochemical competency.
Importantly, these contributions are intended to complement—not replace—existing mechanistic models in plant biology, animal physiology, microbial metabolism, or ecosystem ecology. The framework is designed to operate at a conceptual and comparative level, providing a systems-oriented perspective that can interface with detailed biochemical or ecological models when necessary. In doing so, it expands the theoretical space for cross-disciplinary dialogue and sets the stage for future empirical, computational, and applied extensions in the study of biological performance.
2. Methods
2.1. Conceptual Modeling Approach
This study adopts a theoretical–conceptual modeling approach that aligns with contemporary practices in systems-biology research and scientific theory development (De Regt & Dieks, 2005). Rather than deriving conclusions from direct empirical measurement or experimental data, the framework is constructed through logical synthesis of established principles and cross-disciplinary integration. The modeling process unfolded in several structured stages. First, mass–energy principles shared across a wide range of biological taxa were identified, emphasizing the universal characteristics of living organisms as open, nonequilibrium systems that exchange matter and energy with their surroundings. Second, resource assimilation and physiological losses were formalized using mass-balance expressions, drawing on analogies from ecological energetics, metabolic physiology, and chemical engineering. Third, a competency coefficient was introduced as a conceptual mechanism for capturing biochemical and physiological efficiency, encompassing factors such as enzyme activity, nutrient sufficiency, hormonal regulation, and cellular integrity. Finally, the model was examined for conceptual coherence and alignment with established findings in physiology, plant science, animal bioenergetics, microbial metabolism, and ecological modeling.
Because the purpose of this work is to articulate a generalizable conceptual framework rather than produce numerical predictions, no empirical datasets are analyzed. Instead, the model’s scientific validity is rooted in its consistency with known biological principles, its compatibility with existing theoretical constructs, and its capacity to integrate diverse empirical observations from the literature. This approach allows the framework to operate at a level of abstraction suitable for cross-taxa comparison while avoiding overextension into claims requiring mechanistic or quantitative validation. In this respect, the methodology reflects a theory-building strategy common in systems biology, where conceptual clarity and integrative power are prioritized as precursors to subsequent empirical formalization and computational modeling.
2.2. Biological Assumptions
The development of the proposed framework relies on several biological assumptions that reflect well-established principles across multiple domains of life science. First, it is assumed that all living organisms function as open systems that continuously exchange matter and energy with their environments, a premise grounded in classical thermodynamics and widely accepted in physiology and ecology. Second, the processes of resource uptake and resource loss—denoted as AE (absorbed elements) and TE (transpired or expended elements), respectively—are treated as mass flow rates, allowing assimilation and dissipation to be conceptualized using mass-balance logic. Third, the model assumes that organismal mass (M) scales with metabolic demand, consistent with metabolic scaling theory and empirical observations that larger organisms require greater absolute energy and nutrient throughput. Fourth, it is posited that biochemical competency (CE) modulates the efficiency with which absorbed resources are converted into functional biological outcomes, such as growth, maintenance, reproduction, or stress tolerance. This competency coefficient is understood to encapsulate physiological and biochemical determinants including enzyme activity, nutrient status, hormonal balance, and cellular health. Fifth, while the competency coefficient may vary widely across taxa due to species-specific biochemistry and life-history strategies, it is treated as conceptually general, enabling comparison across organisms without imposing identical mechanistic pathways. Finally, the model explicitly refrains from asserting universal dimensional precision or physical units, acknowledging that the framework yields a comparative biological performance index rather than a physically defined energy measure.
Taken together, these assumptions provide a biologically plausible foundation for conceptual synthesis. They are compatible with established frameworks in bioenergetics, plant physiology, animal nutrition, and metabolic scaling, all of which recognize the central role of mass-energy flux, metabolic efficiency, and organismal size in shaping biological performance..
2.3. Variables and Definitions
For clarity and conceptual consistency, the framework employs a set of defined variables that characterize organismal mass, resource fluxes, and biochemical competency. In this context, M represents the organism’s total mass, expressed in kilograms (kg), and serves as a biologically meaningful scaling factor that reflects absolute metabolic demand. Resource assimilation and dissipation are captured through two mass flow rate variables: AE, denoting the rate of absorbed or assimilated elements (kg·s⁻¹), and TE, denoting the rate of transpired, respired, or excreted elements (kg·s⁻¹). The net outcome of these opposing fluxes over a specified time interval is expressed as Δm, the net mass change (kg) observed over Δt, the time interval measured in seconds (s). Central to the model is the competency coefficient (CE), a dimensionless parameter bounded between 0 and 1 that reflects the organism’s ability to convert absorbed resources into functional biological performance. Unlike purely thermodynamic or mechanistic parameters, CE integrates a range of physiological and biochemical components known to influence metabolic efficiency across taxa.
The competency coefficient aggregates determinants such as enzyme activity, hormonal regulation, vitamin and mineral sufficiency, pigment integrity (including chlorophyll in plants and hemoglobin in animals), as well as cellular and tissue health. Each of these components has been extensively documented in plant and animal physiology as a key modulator of metabolic conversion efficiency, growth potential, and stress tolerance. For example, chlorophyll content directly affects photosynthetic assimilation capacity in plants, while hemoglobin concentration influences oxygen transport efficiency in animals—both outcomes that translate into differences in biological performance. By consolidating these diverse determinants into a single coefficient, the model provides a conceptually tractable means of comparing biochemical competency without requiring species-specific mechanistic detail.
2.4. Derivation of Core Equation
2.4.1. Net Mass Uptake
Mass balance yields:
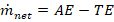
Over a time interval:
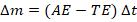
2.4.2. Competency–Ability Equation
We define:
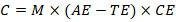
Where C is a biological performance index.
Units are left abstract because is not a physical energy term but a comparative measure.
2.4.3. Time-Integrated Form
Substituting Δm yields:
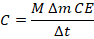
This form aligns with biomass accumulation models and allows longitudinal comparison.
2.5. Physiological Domain Mapping
The model maps to domains as follows:
| Component | Physiological Interpretation |
| AE | Nutrition, photosynthesis, oxygen uptake |
| TE | Respiration, transpiration, excretion |
| M | Structural mass, metabolic scaling |
| CE | Biochemical efficiency & health status |
| C | Functional performance index |
2.6. Scientific Non-Equivalence to Energy Laws
To avoid misinterpretation, this framework explicitly:
- Does not assert a new physical energy law,
- Does not define mechanical energy or joules,
- Does not claim universal dimensional validity.
Instead, it provides a physiology-aligned comparative index compatible with systems-ecology and metabolic theory.
3. Results
Because this is a conceptual paper, results are presented as interpretive scenarios demonstrating applicability across taxa. No numerical predictions are made.
3.1. Plants
3.1.1. Mapping AE and TE
In plants:
- AE corresponds to photosynthetic assimilation + nutrient uptake
- TE corresponds to respiration + transpiration losses
Thus:

Where:
- GPP = gross primary productivity
- R = respiration
- T = transpiration
Empirically, positive net assimilation leads to biomass growth, consistent with plant physiological literature (Taiz et al., 2015).
3.1.2. Competency Coefficient in Plants
Within plants, the competency coefficient (CE) reflects the biochemical and physiological factors that modulate the efficiency with which absorbed resources are converted into biomass, metabolic energy, and structural components. Key contributors to CE include chlorophyll concentration, which directly influences photosynthetic light capture and carbon fixation, and nitrogen availability, which constrains the synthesis of critical enzymes such as Ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco), the most abundant protein in plant leaves and a primary determinant of photosynthetic capacity. Mineral balance also plays an essential role, as elements such as magnesium (Mg) and iron (Fe) are required cofactors for chlorophyll biosynthesis and electron transport, while other micronutrients affect enzyme activation, stomatal functioning, and cellular metabolism. Additional factors such as plant water status and overall tissue integrity further influence biochemical competency by affecting turgor pressure, stomatal conductance, vascular transport, and susceptibility to oxidative damage.
Empirical studies have demonstrated that reductions in any of these components can impair the plant’s physiological efficiency even when external resource availability remains sufficient. For instance, nitrogen deficiency leads to decreased Rubisco content and lowered chlorophyll concentration, thereby reducing carbon assimilation rates and lowering CE despite adequate sunlight and water absorption. Analogously, drought stress may reduce stomatal conductance and impair photosynthetic electron transport, decreasing conversion efficiency independently of light availability. In this way, CE captures the biological reality that resource uptake alone does not guarantee growth or high performance; rather, it is the coordinated function of pigments, enzymes, nutrients, and tissues that determines the degree to which absorbed substrates can be transformed into usable biochemical outputs.
3.2. Animals and Humans
3.2.1. Mapping AE and TE
In animals:
- AE = dietary intake + oxygen uptake
- TE = respiration + excretion + maintenance metabolism
This aligns with nutritional energy balance:

Positive net uptake enables growth, reproduction, and performance; negative values induce catabolism.
3.2.2. Competency Coefficient in Animals
CE corresponds to:
- Enzyme function
- Hormone regulation
- Vitamin/mineral sufficiency
- Immune competency
- Tissue oxygenation
Iron-deficiency anemia, for example, reduces hemoglobin competency, decreasing CE.
In animals, the competency coefficient (CE) encompasses a suite of biochemical andphysiological attributes that determine how efficiently assimilated resources are converted into usable metabolic energy, structural biomass, and functional performance. These attributes include enzyme function, which governs the rate and fidelity of metabolic reactions; hormonal regulation, which mediates growth, metabolism, reproduction, and homeostasis; and vitamin and mineral sufficiency, which ensures proper cofactor availability for enzymatic pathways, mitochondrial respiration, and tissue maintenance. Additional determinants include immune competency, reflecting the organism’s ability to defend against pathogens without excessive energetic cost, and tissue oxygenation, which depends on effective respiratory gas exchange and the transport capacity of oxygen carriers such as hemoglobin. Collectively, these components influence basal metabolic rate, growth efficiency, thermoregulation, reproductive success, and overall physiological resilience.
Importantly, CE captures the biological reality that resource intake does not necessarily translate directly into growth or performance. Animals may consume adequate food and oxygen (high AE), yet exhibit reduced net biological output if internal biochemical systems are compromised. For instance, iron-deficiency anemia reduces hemoglobin concentration and impairs oxygen transport, lowering aerobic metabolic capacity even when dietary intake is sufficient. Under such conditions, CE decreases because metabolic pathways reliant on oxidative phosphorylation become less efficient, forcing greater reliance on anaerobic metabolism or reducing activity and growth altogether. Similar reductions in CE can occur due to micronutrient deficiencies (e.g., vitamin B12, zinc, selenium), endocrine disorders (e.g., hypothyroidism affecting basal metabolic rate), impaired immune function, or chronic inflammation, all of which impose metabolic costs or limit conversion efficiency.
By incorporating these physiological and biochemical determinants into a single dimensionless coefficient, CE provides a conceptually tractable means of comparing metabolic competency across animals without requiring explicit mechanistic modeling of each underlying pathway. This abstraction is particularly useful when evaluating performance across different species, life stages, or environmental contexts where metabolic efficiency varies due to differences in diet quality, physiological condition, or ecological stressors.
3.3. Fish and Aquatic Organisms
Fish and other aquatic organisms operate under physiological constraints that differ markedly from those of terrestrial species, and these constraints directly influence the balance between resource absorption and physiological losses. One major distinction lies in the oxygen acquisition process. Unlike air-breathing animals, fish rely on oxygen diffusion across gill surfaces, a mechanism that is inherently less efficient than pulmonary ventilation due to the substantially lower oxygen content and slower diffusion rates in water. As a result, oxygen uptake (a component of AE) is highly sensitive to the partial pressure of dissolved oxygen, gill surface area, water flow rates, and ventilation–perfusion matching. In hypoxic aquatic environments, oxygen uptake declines, constraining aerobic metabolism and reducing the capacity for biosynthesis, locomotion, and maintenance.
A second defining feature of aquatic physiology is temperature-dependent metabolic rate. As ectotherms, fish exhibit metabolic rates that scale with environmental temperature according to Q10 effects, wherein metabolic reactions accelerate with rising temperature and slow with cooling. Higher temperatures typically increase TE (through elevated respiratory and maintenance costs), while simultaneously raising oxygen demand. If the thermal increase is not matched by sufficient oxygen availability, the result is a mismatch between AE and TE that leads to reduced net resource uptake. Conversely, at lower temperatures metabolic losses decline, but assimilation capacity may also diminish due to reduced digestion efficiency or slowed enzymatic activity.
A third constraint involves nitrogenous waste excretion. Fish primarily excrete nitrogen in the form of ammonia, which is energetically inexpensive to produce but requires adequate water flow for diffusion across gill surfaces. Under conditions of poor water quality or reduced flow, ammonia accumulation can impair gill function and metabolic processes, indirectly reducing AE and increasing physiological stress. Collectively, these features illustrate how aquatic metabolic regulation differs from terrestrial strategies.
These physiological constraints exert strong control over the value of AE − TE, and thus over the competency index C. For instance, fish inhabiting low-oxygen environments (such as warm, eutrophic lakes or poorly aerated aquaculture systems) experience reduced oxygen uptake (lower AE) while simultaneously incurring higher metabolic costs (higher TE), which depresses the net resource balance and lowers overall performance. Similarly, abrupt thermal shifts can alter metabolic costs faster than assimilation capacities can adjust, leading to transient or sustained reductions in C even when food availability is adequate. In this way, the framework accommodates aquatic physiological reality by recognizing that environmental parameters such as temperature, dissolved oxygen, and water chemistry directly modulate both resource assimilation and metabolic expenditures in fish and aquatic organisms.
3.4. Insects
In insects:
- AE = dietary assimilation
- TE = respiration, excretion, molting losses
Molting significantly increases TE and temporarily decreases C due to tissue restructuring.
Insects present another example of how taxon-specific physiology can be interpreted within the competency–ability framework. In these organisms, absorbed elements (AE) correspond primarily to dietary assimilation, encompassing ingestion, digestion, and nutrient absorption through the midgut. The transpired or expended elements (TE) include respiratory gas exchange, excretory losses, and particularly molting-related tissue turnover. Insects undergo periodic molting (ecdysis) as part of their developmental cycle, during which the exoskeleton is shed and replaced. This process imposes substantial metabolic and structural costs, as old cuticular material is degraded and new cuticle is synthesized. Consequently, during molting periods TE increases markedly due to elevated metabolic rates and increased material turnover, leading to a temporary reduction in net resource balance (AE − TE) and thus a transient decrease in the performance index C. Even when food intake remains unchanged, the energetic burden of tissue restructuring and vulnerability to environmental stress can depress CE as well, emphasizing how life-history traits modulate biological performance. Once molting concludes and new tissues stabilize, TE decreases and resource assimilation resumes normal efficiency, illustrating how developmental cycles influence temporal fluctuations in C within insect life histories.
3.5. Microorganisms
For microbes, AE − TE resembles:
- Substrate uptake (AE)
- Maintenance and decay (TE)
This aligns with Monod and chemostat models used in microbial kinetics.
Microorganisms, including bacteria and unicellular eukaryotes, exhibit metabolic dynamics that align closely with mass-balance interpretations of AE − TE. In microbial systems, AE is dominated by substrate uptake, which typically involves transport of dissolved carbon sources, nitrogen compounds, or other nutrients across the cell membrane. Microbial growth kinetics show that substrate assimilation rates depend on external nutrient concentrations, transport system saturation, and enzymatic activity, all of which influence metabolic throughput. Conversely, TE corresponds to maintenance energy requirements, respiratory losses, and decay processes such as lysis or autophagy. These components account for the energetic and material costs required to sustain cellular homeostasis in the absence of net growth. The balance between substrate uptake and maintenance losses determines whether biomass accumulates, remains stable, or declines.
This interpretation aligns closely with established theoretical frameworks in microbial kinetics, particularly Monod models and chemostat dynamics, which describe growth as a function of substrate availability and maintenance energy demands. In these models, microbes exhibit positive growth when substrate uptake exceeds maintenance costs—analogous to AE − TE > 0—and declining biomass when maintenance costs surpass substrate assimilation—analogous to AE − TE < 0. Thus, microbial systems provide a clear example of how net mass balance governs biological performance at the cellular scale. Integrating microbial metabolism into the competency–ability framework underscores its applicability across multiple levels of biological organization, from unicellular organisms to complex multicellular taxa.
4. Discussion
The purpose of this section is to interpret the Universal Life Competency–Ability Framework within the context of established biological theories, evaluate the meaning and implications of the competency coefficient and the composite index , and articulate the advantages, limitations, and prospective research directions associated with this conceptual model. By situating the framework in relation to existing scientific paradigms, we aim to demonstrate both its novelty and its compatibility with accepted principles in physiology, ecology, and systems biology.
4.1. Alignment with Existing Biological Theory
Although the framework was developed conceptually rather than empirically, its components align closely with several well-established theoretical traditions that govern biological energetics, growth, and metabolic scaling. This alignment strengthens the argument that the model is not arbitrary but is instead grounded in widely recognized biological dynamics.
4.1.1. Net Primary Production in Plants
One of the most direct correspondences occurs within plant physiology, specifically in the context of net primary production (NPP). In plants, productivity is commonly expressed as:
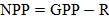
where GPP (gross primary productivity) represents total photosynthetic carbon assimilation, and R (respiration) captures carbon lost through metabolic maintenance and growth processes. This formulation is conceptually equivalent to the expression in the competency framework, where AE represents assimilated carbon and nutrients, and TE represents losses due to respiration, photorespiration, transpiration-driven mass dissipation, and tissue turnover. Thus, NPP provides a direct plant-specific example of how net assimilation drives growth, consistent with the logic that biological performance emerges only when assimilation exceeds losses.
4.1.2. Metabolic Energy Budgets in Animals
Similarly, in animal physiology, energy budgets are frequently expressed in the form:

Here, dietary intake (analogous to AE) must not only cover maintenance and excretory costs (analogous to TE) but also supply surplus energy for growth and reproduction. In this framing, the framework’s performance index can be interpreted as an index of growth and reproduction potential once maintenance and loss requirements have been met. When
is negative, animals enter a catabolic state, reducing performance and eventually compromising survival, which parallels the reductions in
under starvation, metabolic stress, or disease.
4.1.3. Metabolic Scaling Theory
The role of organismal mass M as a scaling factor is further supported by metabolic scaling theory. The seminal work of West, Brown, and Enquist (1997) demonstrated that metabolic rate scales approximately to body mass to the three-quarter power:
This relationship indicates that larger organisms require higher absolute metabolic throughput, consistent with the use of M as a fundamental scaling variable in the competency framework. Although the current model does not explicitly incorporate allometric exponents, treating M as a proportional factor recognizes the empirical reality that metabolic demand increases with organism size.
Collectively, these alignments indicate that the competency framework does not contradict established biological theory; instead, it extends cross-taxonomic abstraction by synthesizing plant-specific, animal-specific, and universal metabolic principles into a shared representation.
4.2. Interpretation of the Competency Coefficient
The competency coefficient (CE) represents one of the most novel elements of the framework. Rather than capturing resource availability or mass flow directly, CE encodes the efficiency of biochemical conversion, integrating physiological determinants that influence how effectively absorbed materials are transformed into usable biological outputs. Conceptually, CE parallels several established metrics across domains:
- In plants, CE is analogous to resource use efficiency (RUE), which represents the ratio of biomass accumulation to resource assimilation (e.g., carbon, nitrogen, or water).
- In animals, CE resembles feed conversion efficiency (FCE), which measures how effectively consumed food contributes to growth or reproduction.
- In microbes, CE aligns with metabolic yield coefficients, which describe how much biomass forms per unit of substrate consumed in batch or chemostat cultures.
By abstracting these analogous constructs into a single coefficient, CE allows biological performance to be compared independent of resource availability, highlighting internal physiological condition rather than external environmental supply. This distinction is critical, as organisms experiencing identical resource inputs may exhibit dramatically different performance due to disease, deficiency, hormonal imbalance, or tissue damage. Thus, CE captures the idea that biological “competency” is not merely a function of supply but of the capacity to utilize supply.
4.3. Biological Meaning of
The composite index should not be interpreted as a physical quantity such as joules, watts, or mechanical energy. Instead, it provides a functional biological performance index, integrating mass balance and conversion efficiency into a single interpretable measure. As such,
reflects emergent organismal traits including:
- Growth potential, as surplus mass-energy supports biosynthesis.
- Physiological vitality, reflecting metabolic and biochemical capacity.
- Stress resilience, indicating robustness under environmental perturbation.
- Reproductive capacity, as reproduction typically requires positive net resource balance and high biochemical competency.
Because is dimensionally abstract, it is especially suited for comparative, diagnostic, and conceptual applications rather than quantitative bioenergetic modeling.
4.4. Stress, Deficiency, and Disease Effects
Environmental stressors typically modulate by reducing AE, increasing TE, decreasing CE, or some combination thereof. Table-like trends include:
| Stressor | AE | TE | CE |
| Drought in plants | ↓ | ↑ | ↓ |
| Starvation in animals | ↓ | ↑ | ↓ |
| Mineral deficiency | — | — | ↓ |
| Thermal stress | ↓ | ↑ | ↓ |
| Disease | ↓ | ↑ | ↓ |
Under such conditions:
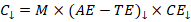
This illustrates that even without direct changes in environmental resources, physiological or biochemical damage can sharply reduce performance by lowering CE.
4.5. Advantages of the Framework
The competency–ability framework offers several conceptual advantages. First, it provides taxonomic universality, enabling discussion of plants, animals, microbes, and insects using common terminology. Second, it affords conceptual clarity by distinguishing resource availability, physiological losses, and conversion efficiency. Third, it maintains mass-balance coherence, aligning with established principles in ecology and bioenergetics. Fourth, it demonstrates compatibility with existing literature, as shown in Section 4.1. Finally, it retains non-mechanistic flexibility, facilitating cross-disciplinary interpretation without requiring detailed mechanistic modeling.
4.6. Limitations
Despite its utility, the framework has limitations that merit acknowledgment. It is currently not empirically calibrated, meaning numerical values for C lack quantitative grounding. CE remains a qualitative construct requiring operational definitions for measurement, and the model does not incorporate allometric exponents, which are essential for scaling metabolic rates precisely. Furthermore, the framework is not designed for predictive precision, limiting its utility in simulations or engineering applications. It also does not replace domain-specific models, which remain indispensable for mechanistic insight. These limitations suggest that the framework should be interpreted as a conceptual scaffold rather than a predictive model.
4.7. Future Research Directions
The conceptual nature of the model invites extensive avenues for empirical and computational development. Future work may focus on operationalizing CE through measurable biomarkers such as chlorophyll content, photosynthetic enzyme activity, blood oxygen saturation, hormonal panels, micronutrient concentrations, or immune indices. Integration of metabolic scaling laws could refine the role of mass, for example by incorporating or surface-area scaling terms. Computational modeling approaches—such as agent-based models, differential equation systems, or network simulations—could translate conceptual structure into dynamic prediction. Application domains are diverse: in agriculture, the framework could support crop stress indexing or livestock productivity assessment; in ecology, it may inform studies of climate stress resilience or invasive species performance; and in biomedicine, it could aid in analyzing metabolic disorders or nutritional deficiencies. Collectively, these directions underscore the framework’s potential for interdisciplinary extension.
5. Conclusion
This paper introduced the Universal Life Competency–Ability Framework, a conceptual systems-biology model that integrates organismal mass, net resource uptake, and biochemical competency into a biologically meaningful performance index. The model does not propose new physical laws but instead synthesizes established principles from physiology, ecological energetics, and metabolic theory into a unified comparative structure.
The resulting expression:
provides insight into how resource assimilation, physiological loss, and biochemical efficiency interact to shape growth, vitality, and resilience across diverse life forms. Conceptual analysis demonstrates alignment with classical plant and animal physiology as well as metabolic scaling and ecological production models.
The principal contribution of this work is to articulate a taxonomically general, mass-balance-grounded perspective on biological performance without overclaiming quantitative precision. Future research may focus on empirical calibration, incorporation of metabolic scaling exponents, and development of domain-specific applications in biomedicine, agriculture, ecological modeling, and bioengineering.
In conclusion, the Universal Life Competency–Ability Framework offers a scientifically defensible conceptual tool for interpreting biological performance within and across taxa, complementing existing mechanistic models and advancing systems-level understanding of life processes.
References
World Health Organization. (2022). Global competency and outcomes framework for universal health coverage. World Health Organization.
Gómez-Rey, P., Barbera, E., Fernández-Navarro, F., Zhang, J., & Teixeira, A. M. (2021). Development and validation of a life skills evaluation tool for online learning based on the framework of the capability approach. Educational Technology Research and Development, 69(6), 3029-3049.
Clemmons, A. W., Timbrook, J., Herron, J. C., & Crowe, A. J. (2020). BioSkills Guide: Development and national validation of a tool for interpreting the Vision and Change core competencies. CBE—Life Sciences Education, 19(4), ar53.
Martimianakis, M. A. T., & Hafferty, F. W. (2013). The world as the new local clinic: a critical analysis of three discourses of global medical competency. Social Science & Medicine, 87, 31-38.
Masaev, S. N., Dorrer, G. A., Minkin, A. N., Bogdanov, A. V., & Salal, Y. K. (2020, November). Assessment of the application of the Universal Competencies. In Journal of Physics: Conference Series (Vol. 1691, No. 1, p. 012020). IOP Publishing.
Ala-Mutka, K. (2011). Mapping digital competence: Towards a conceptual understanding. Sevilla: Institute for Prospective Technological Studies, 7-60.
Shambare, B., & Simuja, C. (2024). Mapping TPACK Competency among Life Sciences Teachers in Rural and Marginalised Secondary Schools: A Descriptive Analysis. International Journal of Technology in Education and Science, 8(4), 522-541.
Qiao, C., Chen, Y., Guo, Q., & Yu, Y. (2024). Understanding science data literacy: A conceptual framework and assessment tool for college students majoring in STEM. International Journal of STEM Education, 11(1), 25.
Zarcadoolas, C., Pleasant, A., & Greer, D. S. (2006). Advancing health literacy: A framework for understanding and action. John Wiley & Sons.
Frye, K. E., Boss, D. L., Anthony, C. J., Du, H., & Xing, W. (2024). Content analysis of the CASEL framework using K–12 state SEL standards. School Psychology Review, 53(3), 208-222.
Andino-González, P., Vega-Muñoz, A., Salazar-Sepúlveda, G., Contreras-Barraza, N., Lay, N., & Gil-Marín, M. (2025). Systematic review of studies using confirmatory factor analysis for measuring management skills in sustainable organizational development. Sustainability, 17(6), 2373.
De Regt, H. W., & Dieks, D. (2005). A contextual approach to scientific understanding. Synthese, 144(1), 137-170.
Green, F. (2013). Skills and skilled work: an economic and social analysis. Oxford University Press, USA.

You must be logged in to post a comment.