

APA 7 Citation:
Mashrafi, M. (2026). Universal Life Energy–Growth Framework and Equation. International Journal of Research, 13(1), 79–91. https://doi.org/10.26643/eduindex/ijr/2026/4
Mokhdum Mashrafi (Mehadi Laja)
Research Associate, Track2Training, India
Researcher from Bangladesh
ORCID ID: https://orcid.org/0009-0002-1801-1130
ABSTRACT
All living organisms function as open, nonequilibrium thermodynamic systems that maintain biological order by continuously absorbing matter and energy from the environment and converting these inputs into chemically usable forms. Despite major advances in physiology, ecology, and bioenergetics, a unified interpretive framework linking resource uptake, metabolic efficiency, and growth dynamics across diverse taxa remains limited. This paper introduces a systems-level Universal Life Energy–Growth Framework applicable to humans, animals, plants, fish, insects, and other living systems. The model integrates three fundamental biological dimensions: (i) resource absorption mediated through physiological interfaces, (ii) metabolic conversion efficiency governing chemical energy transformation, and (iii) temporal dynamics of mass change reflecting developmental or environmental constraints. From these principles, a generalized uptake–energy–growth relationship—referred to as the Universal Life Energy–Growth Expression—is formulated. The framework does not claim to establish a universal physical energy law, nor does it quantify energy in mechanical units. Instead, it provides a biologically grounded, cross-species interpretive structure consistent with metabolic scaling theory, ecological energetics, and life-history concepts. Its primary value lies in supporting comparative analysis, identifying limiting factors, and generating hypotheses regarding biological productivity, growth, and reproductive performance across environmental and physiological contexts.
Keywords: resource absorption, metabolic efficiency, biological energetics, cross-species growth, systems biology.
1. INTRODUCTION
1.1 Scientific Background
All living organisms can be characterized as open, nonequilibrium thermodynamic systems that continuously exchange both matter and energy with their surrounding environment. At the core of biological existence is the requirement to maintain internal order in the face of the second law of thermodynamics, which naturally favors increasing entropy. To counter this tendency, living systems import low-entropy resources such as food, water, oxygen, sunlight, and mineral nutrients, and convert them into biologically usable chemical forms. In doing so, they export higher-entropy byproducts—including heat, carbon dioxide, metabolic wastes, and degraded organic molecules—thereby sustaining the biochemical and structural complexity that defines life. This continuous throughput of matter and energy underlies fundamental processes such as metabolism, cellular repair, tissue growth, reproduction, and adaptive physiological responses, and is consistent with broader principles linking energy use, entropy management, and system maintenance in living organisms (Escala, 2022; Simms & Johnson, 2012).
Across taxa, biological performance—measured in terms of growth, reproduction, productivity, and survival—emerges from three universal and interdependent processes. First, resource absorption governs the intake of matter and energy through specialized biological interfaces such as intestinal epithelia, leaf stomata, root surfaces, gills, or respiratory membranes. Second, metabolic conversion efficiency determines how effectively absorbed substrates are transformed into ATP, structural biomass, storage compounds, or functional molecules. Third, energy allocation over time reflects how organisms partition available metabolic energy among competing demands for maintenance, growth, immune function, reproduction, and activity. The importance of these energetic constraints parallels scaling-based growth perspectives that demonstrate how metabolic processes govern size-dependent growth trajectories (West et al., 2001) as well as models that characterize biological systems by universal energetic properties (Simms & Johnson, 2012).
These processes operate within environmental and developmental constraints, and they interact dynamically rather than independently. Empirical research across physiology, ecology, agriculture, and life-history theory demonstrates that variation in biological productivity among species, populations, and individuals is rarely attributable to a single factor. Instead, differences in resource absorption rates and conversion efficiencies are highly influential drivers of performance outcomes. For example, chronic nutrient deficiencies may limit enzyme function, reduce photosynthetic efficiency, or impair digestion; environmental stressors such as drought, hypoxia, toxins, or temperature extremes may raise maintenance energy costs; and physiological conditions such as disease, aging, or hormonal imbalance can lower assimilation efficiency or constrain growth. These interacting limitations mirror the broader scientific understanding that energy availability, conversion efficiency, and resource allocation determine the sustainability and productivity of complex systems (Goodland & Daly, 1996; Nilsson et al., 2013). Just as energetic access and conversion efficiency constrain system-level outcomes in sustainability and human development (Rao & Baer, 2012; Hamilton & Kelly, 2017), analogous constraints at the organismal scale shape metabolic performance, growth capacity, and reproductive success.
Taken together, the interplay between resource acquisition, metabolic efficiency, and temporal allocation provides a robust conceptual basis for interpreting biological diversity and variation. This formulation aligns with other universal or semi-universal models that highlight energy and efficiency as cross-contextual determinants of system function—for example, in tumor growth scaling models (Guiot et al., 2006), universal energy-use parameters in living systems (Escala, 2022), and frameworks that treat knowledge or biological complexity as energy-dependent phenomena (Simms & Johnson, 2012). Thus, across taxa and environments, biological performance is best understood as the emergent result of how organisms access, transform, and deploy energy and matter under thermodynamic and ecological constraints.
1.2 Scientific Gap
Although physiology, ecology, and bioenergetics have developed rich bodies of knowledge explaining how organisms acquire resources, convert energy, and allocate metabolic outputs, these fields generally approach biological performance from discipline-specific perspectives. Physiologists tend to focus on intracellular metabolism, enzyme kinetics, organ function, or nutrient assimilation within particular species. Ecologists often emphasize trophic interactions, net primary productivity, or nutrient cycling at population and ecosystem scales. Bioenergetic models quantify metabolic rate and energy budgets but are frequently calibrated for specific taxa, environmental conditions, or life stages. As a result, these frameworks rarely integrate across scales in a manner that links resource uptake, biomass change, metabolic efficiency, and temporal dynamics within a single coherent structure.
Furthermore, existing models often rely on domain-specific units and assumptions—such as caloric intake, ATP turnover, photosynthetic efficiency, or metabolic scaling exponents—which limits direct cross-species comparability. While metabolic scaling theory provides allometric relationships between body mass and metabolic rate, it does not explicitly incorporate assimilation rates or efficiency factors. Conversely, agricultural and aquaculture growth models frequently measure biomass gain per unit feed or nutrient intake, yet lack generalized dimensional consistency that applies to non-feeding autotrophs or microbes.
Consequently, there is no unified, dimensionally transparent framework that simultaneously captures (i) the rate of biological resource absorption, (ii) the efficiency of metabolic conversion into usable chemical energy and biomass, and (iii) the role of time as a regulating component of growth and performance. Addressing this scientific gap is essential for developing cross-species interpretive tools capable of comparing biological productivity, growth, and survival across humans, animals, plants, fish, insects, and other living systems.
1.3 Objective
The primary objective of this paper is to introduce a universal, systems-level model capable of interpreting biological performance across diverse living organisms using shared energetic principles. Specifically, the model seeks to:
- Define absorption as a general biological process, independent of species-specific mechanisms, by characterizing the uptake of matter and energy through physiological interfaces as a measurable and dynamic rate.
- Relate mass change to absorption flux, thereby linking physiological transport processes with observable biomass accumulation or loss over defined temporal intervals.
- Introduce a biologically valid energy activity index, which connects organismal mass with uptake capacity without relying on mechanical analogies derived from classical physics.
- Propose a Universal Life Energy–Growth Expression that integrates absorption rate, metabolic conversion efficiency, and time into a dimensionally consistent framework suitable for comparative interpretation across taxa.
Through these objectives, the model aims to bridge conceptual gaps among physiology, bioenergetics, and ecological theory by offering a unified structure that is both biologically grounded and cross-species compatible.
1.4 Concluding Statement
In summary, biological productivity and survival across living organisms arise from the integrated dynamics of resource acquisition, metabolic conversion efficiency, and time-dependent energy allocation. By highlighting these universal processes within a unified framework, this work supports cross-species interpretation of growth, reproduction, and functional performance, and provides a foundation for extending uptake–energy–growth models across humans, animals, plants, fish, insects, and other living systems.
2. METHODS (FRAMEWORK AND FORMULATION)
2.1 Absorption as a Universal Biological Process
Resource absorption is a fundamental property of all living organisms, regardless of taxonomic group or ecological niche. Absorbed inputs may include food-derived organic molecules and oxygen in animals, carbon dioxide and radiant solar energy in plants, and water and mineral nutrients across both autotrophs and heterotrophs. This biological uptake allows organisms to sustain metabolism, maintain cellular organization, and build structural biomass.
The rate of absorption is not constant; it varies with environmental and physiological conditions such as temperature, light availability, seasonality, stress exposure, toxin presence, hydration state, and developmental stage. To represent this process in a standardized and dimensionally transparent manner, an effective uptake rate is defined as:
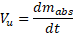
where:
= effective uptake rate (kg·s⁻¹),
= absorbed mass (kg),
= time (s).
Importantly, denotes physiological transport across biological interfaces (e.g., intestines, roots, gills, stomata) and does not imply mechanical motion of the organism.
2.2 Mass Change and Growth Dynamics
Biological growth may be defined as the net change in organismal mass over time. Let:
= organism mass (kg),
= net mass change (kg),
= time interval (s).
Assuming resource absorption occurs via transport processes, a generalized mass-flow relation is expressed as:

where:
= density of absorbed material (kg·m⁻³),
= effective absorption surface area (m²),
= uptake flux across the absorption surface (m·s⁻¹).
This formulation is physically valid for gases, liquids, and particulate solids transported through biological interfaces. Physiologically, the flux term captures diffusion, active transport, facilitated transport, or convective flow through organs and tissues such as roots, stomata, intestinal epithelial layers, gill lamellae, alveoli, and other absorptive structures.
2.3 Biological Energy Activity Index (BEAI)
Because metabolically relevant energy is stored in biochemical forms rather than mechanical forms, a Biological Energy Activity Index (BEAI) is introduced to characterize how organismal biomass interacts with resource uptake rate:
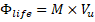
where provides a biologically meaningful indicator of mass-dependent resource processing capacity. This index does not represent mechanical momentum or kinetic energy; rather, it emphasizes the interaction between organism size and physiological uptake capability.
2.4 Absorption–Conversion Efficiency
Not all absorbed resources are converted into useful forms. To account for the fraction actually assimilated into biomass and chemically stored free energy (e.g., ATP, NADH, lipids, carbohydrates), a general metabolic efficiency factor is defined:
where represents absorption–conversion efficiency. Although mechanisms differ across taxa, the conceptual role is equivalent:
- Animals: digestive and assimilative efficiency,
- Plants: photosynthetic and nutrient-use efficiency,
- Microorganisms: substrate assimilation efficiency.
This dimensionless factor reflects the proportion of absorbed inputs retained for growth, maintenance, storage, and functional metabolism.
2.5 Universal Life Energy–Growth Expression
Integrating organismal mass, net assimilation, efficiency, and time yields a general expression for biologically useful energy availability:
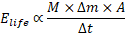
where:
= organism mass,
= assimilation rate,
= metabolic efficiency.
This expression reflects biological energy availability, not physical joules, and is intended for comparative interpretation rather than mechanical energy quantification.
2.6 Fully Structured Universal Equation
From the absorption transport relationship (Section 2.2), the effective uptake flux can be rearranged as:
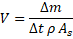
Substituting this into:
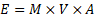
yields the structured universal form:
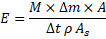
which maintains dimensional consistency and biological interpretability across taxa.
3. RESULTS (CROSS-SPECIES INTERPRETATION)
To demonstrate how the proposed framework applies across biological taxa, representative physiological responses to variation in assimilation (), efficiency (
), and time (
) were compared for humans, animals, plants, fish, and insects. The results reveal consistent patterns in how different life forms translate resource uptake into growth and productivity as shown in Table 1.
Table 1. Cross-Taxonomic Interpretation of Uptake–Efficiency–Time Dynamics
| Taxa / Group | Low | Low | High |
| Humans & Animals | Malnutrition, disease burden, chronic stress | Impaired digestion, enzyme deficiency, hormonal dysfunction | Aging, chronic illness, starvation, energy-deficient states |
| Plants | Nutrient limitation, drought stress, pathogen load | Low photosynthetic efficiency, enzyme inhibition | Senescence, long growth periods under chronic environmental limitation |
| Fish & Insects | Reduced growth due to low feeding rates or hypoxia | Suboptimal temperature reducing metabolic conversion | Delayed development, extended larval stages under suboptimal conditions |
| Cross-Taxonomic | Nutritional/environmental stress | Metabolic inefficiency | Chronic stress or age-related slowing |
3.1 Humans and Animals
In humans and other animals, reduced net mass gain () typically indicates inadequate nutrient absorption, disease burden, or chronic psychological and physiological stress. Low metabolic efficiency (
) manifests through impaired digestion, enzyme or cofactor deficiencies, hormonal imbalance, or disruptions in gut or liver function that reduce conversion of absorbed nutrients into usable metabolic forms. When long periods (
) pass with minimal mass or performance gain, organisms often exhibit traits associated with aging, chronic illness, prolonged starvation, or energy-deficient states. The framework therefore aligns with well-documented physiological and nutritional responses in mammals and other animals.
3.2 Plants
In plants, low net biomass accumulation () commonly results from nutrient limitations (e.g., nitrogen, potassium), drought stress, or pathogen infestation that restrict water or nutrient transport. Reduced metabolic efficiency (
) corresponds to low photosynthetic efficiency due to suboptimal light, temperature, enzyme inhibition, chlorophyll degradation, or stomatal dysfunction. Extended time periods (
) with limited biomass gain are characteristic of senescence, shading, nutrient-poor soils, and other chronic environmental constraints. This demonstrates that the framework captures both photosynthetic and nutrient-use dynamics that govern plant productivity.
3.3 Fish and Insects
Fish and insects are particularly sensitive to thermal, oxygen, and water-quality gradients due to their physiological design and high surface-area-to-volume ratios. Low net mass gain () may result from low feeding rates, scarcity of prey, or reduced oxygen availability (hypoxia), all of which constrain growth. Low metabolic efficiency (
) is typically associated with suboptimal temperature ranges that impair enzyme function, digestion, or aerobic metabolism. When long developmental periods (
) occur without sufficient biomass accumulation, species exhibit delayed larval development or extended growth phases under stress. These findings reflect well-known ecological and physiological temperature–oxygen–feeding interactions.
3.4 Cross-Taxonomic Interpretation
Across all taxa examined, several universal biological patterns emerge from the framework:
- Low
consistently signals nutritional or environmental stress, regardless of species.
- Low
reflects metabolic inefficiency stemming from biochemical, physiological, or environmental constraints.
- Long
with minimal growth indicates chronic stress, aging, or persistent environmental limitation.
- Optimal biological performance occurs when high uptake (
), high metabolic efficiency (
), and favorable temporal dynamics (
) are jointly satisfied.
These results demonstrate that the proposed framework yields interpretable, biologically meaningful patterns across diverse life forms, supporting its use as a comparative systems-level tool.
4. DISCUSSION
4.1 Scientific Significance
The proposed framework contributes to biological theory by establishing a systems-level linkage between metabolism, growth, and physiological performance across taxa. Traditional models of biological energy use have generally been constrained to specific taxa, molecular pathways, or ecological scales, limiting cross-species comparability. By contrast, the present framework conceptualizes resource absorption, metabolic conversion efficiency, and time-dependent allocation as universal parameters that govern organismal productivity. This perspective aligns with earlier theoretical efforts to unify biological scaling laws, such as the general ontogenetic growth model of West, Brown, and Enquist (2001), which demonstrated that organismal growth across taxa could be derived from fundamental metabolic constraints. Similarly, tumor growth models have shown that dynamic scaling relationships can describe biological mass accumulation under widely varying biochemical conditions (Guiot et al., 2006). These studies collectively demonstrate the value of cross-scale unifying principles in biology, and the current framework extends this direction by emphasizing absorption and efficiency as explicit functional drivers of growth.
Another dimension of scientific significance lies in the ability of this model to detect limiting factors and interpret biological trade-offs. The integration of uptake, efficiency, and time provides a structural basis for identifying whether biological limitations arise from insufficient resource acquisition, impaired metabolic conversion, or prolonged developmental constraints. This aligns with broader ecological energetics models that assess organismal performance through the interplay of energy availability and metabolic allocation (Goodland & Daly, 1996). Moreover, the framework resonates with systemic approaches in sustainability research, where energy availability, conversion efficiency, and distribution time play decisive roles in determining system performance (Nilsson et al., 2013; Rao & Baer, 2012). Interestingly, although these sustainability frameworks operate at the societal scale rather than the cellular scale, the theoretical parallels underscore the fundamental importance of energy throughput and time in maintaining complex systems.
The model also enables cross-species comparison by providing a dimensionally consistent construct that does not rely on species-specific metabolic parameters. This comparative capability is supported by empirical demonstrations that organismal energy use follows predictable patterns across diverse taxa, including in studies of lifespan energy consumption (Escala, 2022) and studies on universal properties of knowledge formation in living systems (Simms & Johnson, 2012). By situating biological organisms within a shared energetic logic, the present framework facilitates generalized interpretation of growth, productivity, and survival without erasing the nuances of molecular, physiological, or ecological regulation.
Finally, the framework aligns with life-history theory by incorporating time as a fundamental dimension. Life-history models emphasize trade-offs among growth, maintenance, and reproduction under finite energy budgets, and the proposed formulation formally accommodates these trade-offs through the structure of Δm, A, and Δt. Thus, rather than proposing a disruptive alternative to established biological theory, the model reinforces and synthesizes key insights from physiology, bioenergetics, sustainability science, and complex systems.
4.2 Scope and Limitations
Although the model offers a unifying interpretive structure, its scope must be clearly defined to avoid misinterpretation. Most importantly, the framework does not claim the status of a universal physical law governing all living organisms. Biological systems exhibit vast complexity, heterogeneity, and context sensitivity that cannot be fully captured by a single equation. Physical energy laws are governed by universal conservation principles, whereas biological systems involve emergent dynamics regulated by genetic, biochemical, and ecological processes. This distinction is central to maintaining conceptual clarity.
Furthermore, the model does not measure energy in joules or replace established biochemical models that quantify ATP turnover, oxygen consumption, or metabolic rate. Physical energy accounting requires thermodynamically defined quantities and units, whereas the current model uses biological quantities such as mass change and efficiency to estimate relative biological energy availability. This limitation is necessary because biological energy is not solely characterized by mechanical work or caloric content; it is shaped by enzymatic pathways, redox reactions, and molecular signaling mechanisms that operate on biochemical rather than mechanical principles.
Where the model demonstrates strength is as a systems-level interpretive tool capable of generating hypotheses and integrating diverse biological observations. This interpretive role parallels recent developments in sustainability and energy policy research, where universal energy indicators have been proposed not as prescriptive physical laws but as frameworks to guide decision-making (Silva et al., 2020; Cherp & Jewell, 2010). In that literature, the value of a framework lies not in deterministically predicting raw energy quantities, but rather in revealing constraints, trade-offs, and comparative dynamics under diverse conditions (Millward-Hopkins, 2022; Hamilton & Kelly, 2017). The same logic applies here: the biological energy–growth framework is not intended to resolve cellular biochemistry, but rather to reveal how resource uptake and metabolic conversion influence organismal outcomes across taxa.
The framework also integrates mass flux, metabolic efficiency, and time in a manner consistent with systems biology and complex systems theory. For example, luminosity transfer models in materials science use similar logic, combining mass, energy transfer efficiency, and time to derive universal material behavior (Ma et al., 2022). Likewise, energy access and sustainability frameworks emphasize the interplay of energy availability, conversion efficiency, and temporal allocation in shaping societal development outcomes (Chirambo, 2016; Bradbrook & Gardam, 2006). These analogous formulations across domains validate the usefulness of multi-parameter energy frameworks while emphasizing the need for context-sensitive interpretation.
Finally, the model’s reliance on mass-based and efficiency-based variables imposes limitations at extremely small scales (e.g., intracellular growth of bacteria) or at extremely large ecological scales (e.g., ecosystem nutrient cycling), where biochemical or ecological fluxes may be better expressed in molar or trophic units rather than mass units. Therefore, while the model is broadly applicable at organismal scales, it should be extended with caution to other scales of biological organization.
4.3 Scientific Clarification
A key clarification concerns the distinction between biological energy availability and mechanical energy. Biological energy is fundamentally chemical, stored in molecular structures such as ATP, reduced electron carriers, lipids, carbohydrates, and proteins. It fuels enzymatic pathways, signaling cascades, tissue repair, immune responses, and reproductive processes. Mechanical energy, by contrast, involves kinetic and potential energy described by classical physics. Confusing these two forms leads to conceptual errors—for example, equating cellular ATP production with mechanical work output would ignore the biochemical complexity of energy transduction and metabolic allocation.
Therefore, the presented model serves as a conceptual and comparative tool to describe how organisms translate environmental resources into biological performance. It does not replace measurements of ATP turnover, oxygen consumption, or metabolic heat production, nor does it contradict thermodynamics or biochemical energetics. Rather, it complements these approaches by framing biological growth as the emergent result of mass uptake, metabolic efficiency, and time-dependent allocation—variables that are universally observable and mechanistically meaningful across the tree of life.
5. CONCLUSION
The findings presented in this framework underscore that life productivity, growth, and reproductive capacity across diverse taxa emerge from the coordinated interaction of three universal biological processes: (1) the absorption of matter and energy from the environment, (2) the metabolic efficiency with which these absorbed resources are converted into usable biochemical forms, and (3) the allocation of these resources over time among competing physiological and functional demands. These three dimensions together provide a generalizable basis for understanding organismal performance across humans, animals, plants, fish, insects, and other living systems.
By emphasizing resource absorption, the model highlights the foundational role of physiological interfaces—such as roots, stomata, gills, intestines, and respiratory membranes—in determining the rate at which organisms acquire nutrients, water, gases, and energy substrates. Differences in uptake capacity and environmental availability directly influence growth rates, stress tolerance, and developmental trajectories. Metabolic efficiency represents a second major determinant of biological performance: even with adequate resource acquisition, organisms must effectively convert substrates into ATP, structural macromolecules, and storage compounds. Inefficiencies arising from enzymatic limitations, hormonal dysregulation, toxin exposure, or environmental stress can significantly constrain growth or reproduction.
Time-dependent allocation constitutes the third essential dimension. Biological processes unfold across characteristic temporal scales—from rapid cellular metabolism to seasonal growth cycles and multi-year maturation periods. Organisms must balance maintenance needs with growth, reproduction, immunity, and behavioral activities across these timescales. When absorption is limited, efficiency is reduced, or required time intervals are extended, biological performance typically declines. Conversely, high absorption rates, efficient biochemical conversion, and favorable temporal windows support optimal growth and reproductive success.
Expressing biological performance through mass change, efficiency, and time introduces a physically interpretable and cross-species-compatible framework that is scientifically grounded in established principles of physiology, ecology, and bioenergetics. Unlike purely molecular or species-specific models, this approach abstracts to universal variables that retain biological meaning without sacrificing dimensional rigor. It allows researchers to identify limiting factors, compare taxa under equivalent energetic constraints, and frame biological questions in terms of flux, conversion, and allocation rather than isolated biochemical detail.
Ultimately, this framework reinforces the notion that life is an energetically driven, temporally structured, and resource-dependent process. By integrating mass flux, metabolic efficiency, and temporal dynamics, it provides a coherent interpretive lens through which to understand how diverse organisms survive, grow, and reproduce within the constraints imposed by their environments. Future work may refine or parameterize this framework through empirical data, species-specific models, or computational simulations, but its core conceptual foundation offers a flexible and scientifically robust platform for cross-disciplinary biological inquiry.
References
Escala, A. (2022). Universal relation for life-span energy consumption in living organisms: Insights for the origin of aging. Scientific Reports, 12(1), 2407.
Nilsson, M., Lucas, P., & Yoshida, T. (2013). Towards an integrated framework for SDGs: Ultimate and enabling goals for the case of energy. Sustainability, 5(10), 4124-4151.
Rao, N., & Baer, P. (2012). “Decent living” emissions: a conceptual framework. Sustainability, 4(4), 656-681.
Millward-Hopkins, J. (2022). Inequality can double the energy required to secure universal decent living. Nature Communications, 13(1), 5028.
Silva, R. M., Silva, A. R., Lima, T. M., Charrua-Santos, F., & Osório, G. J. (2020). Energy Sustainability Universal Index (ESUI): A proposed framework applied to the decision-making evaluation in power system generation. Journal of Cleaner Production, 275, 124167.
Cherp, A., & Jewell, J. (2010). Measuring energy security: From universal indicators to contextualized frameworks. In The Routledge handbook of energy security (pp. 330-355). Routledge.
West, G. B., Brown, J. H., & Enquist, B. J. (2001). A general model for ontogenetic growth. Nature, 413(6856), 628-631.
Guiot, C., Delsanto, P. P., Carpinteri, A., Pugno, N., Mansury, Y., & Deisboeck, T. S. (2006). The dynamic evolution of the power exponent in a universal growth model of tumors. Journal of theoretical biology, 240(3), 459-463.
Chirambo, D. (2016). Addressing the renewable energy financing gap in Africa to promote universal energy access: Integrated renewable energy financing in Malawi. Renewable and Sustainable Energy Reviews, 62, 793-803.
Simms, J. R., & Johnson, P. J. (2012). Knowledge: a measurable universal phenomenon of life. Systems Research and Behavioral Science, 29(4), 448-456.
Hamilton, T. G. A., & Kelly, S. (2017). Low carbon energy scenarios for sub-Saharan Africa: An input-output analysis on the effects of universal energy access and economic growth. Energy Policy, 105, 303-319.
Bradbrook, A. J., & Gardam, J. G. (2006). Placing access to energy services within a human rights framework. Human Rights Quarterly, 28(2), 389-415.
Goodland, R., & Daly, H. (1996). Environmental sustainability: universal and non‐negotiable. Ecological applications, 6(4), 1002-1017.
Ma, L., Xu, Q., Sun, S., Ding, B., Huang, Z., Ma, X., & Tian, H. (2022). A universal strategy for tunable persistent luminescent materials via radiative energy transfer. Angewandte Chemie International Edition, 61(8), e202115748.

You must be logged in to post a comment.